Diplópode
Diplopoda | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
Ocorrência: Silúrico tardio-actual (há 428-0 milhões de anos) 428–0 Ma PreЄ Є O S D C P T J K Pg N | |||||||||||
 Vários diplópodes (sem escala) | |||||||||||
Classificação científica | |||||||||||
| |||||||||||
Diversidade | |||||||||||
| 16 ordens, c. 12 000 espécies | |||||||||||
| Subclasses | |||||||||||
| |||||||||||
Os diplópodes[1] (Diplopoda) são uma classe do subfilo Myriapoda, vulgarmente conhecidos como piolhos-de-cobra. Vivem em ambientes húmidos, com pouca luminosidade e com material orgânico disponível para a alimentação,[2][3] podendo ser carniceiros e parasitas de plantas. Outros nomes pelos quais são conhecidos incluem embuá, gongolo e variantes.[3]
Os diplópodes são um grupo de artrópodes que se caracterizam por ter dois pares de patas na maioria dos seus segmentos corporais e que se classificam taxonomicamente como uma classe. Cada segmento com dois pares de patas é o resultado da fusão de dois segmentos simples. A maioria dos diplópodes tem corpos cilíndricos muito alongados ou corpos aplanados com mais de 20 segmentos, enquanto que os milípedes Oniscomorpha são mais curtos e podem enrolar-se formando uma bola. Apesar de em muitas línguas se lhes dar o nome de "milípedes" ("mil-pés"), nenhuma espécie tem realmente mil patas; o registo que se tem é da espécie de 750 patas Illacme plenipes. Existem cerca de 12 000 espécies designadas de diplópodes,[4] classificadas em 16 ordens e cerca de 140 famílias, o que faz com que sejam a maior classe de miriápodes, superando mesmo os quilópodes.
A maioria dos diplópodes são detritívoros, com movimentos muito lentos, que comem folhas podres caídas e outras matérias vegetais. Alguns comem fungos ou sugam fluidos de plantas, e uma pequena minoria são predadores. Os diplópodes são geralmente inofensivos para os humanos, apesar de alguns poderem transformar-se em verdadeiras pragas nas casas ou nos jardins, principalmente em estufas, onde podem causar graves danos aos rebentos de plantas recém-nascidas. A maioria defende-se segregando diversos compostos químicos através de poros que têm pelo corpo, embora os pequenos Polyxenida estejam munidos de penachos de cerdas que se desprendem. A reprodução na maioria das espécies é levada a cabo por transferência de pacotes de esperma às fêmeas, que se realiza por meio de patas modificadas do macho chamadas gonóporos.
Apareceram pela primeira vez no período Silúrico, sendo uns dos animais terrestres mais antigos de que se tenha conhecimento. Alguns membros de grupos pré-históricos chegaram a alcançar os 2 m de comprimento, mas as maiores espécies modernas apresentam um comprimento máximo de entre 27 e 38 cm, e a maior de todas acredita-se que seja o milípede gigante africano (Archispirostreptus gigas).
Entre os miriápodes, os diplópodes foram considerados tradicionalmente mais próximos aos pequenos paurópodes, apesar de alguns estudos moleculares porem em causa esta relação. Os diplópodes podem distinguir-se das semelhantes mas apenas distantemente relacionadas centopeias (classe Chilopoda), mas estas últimas se movem rapidamente, são carnívoras e têm só um par de patas em cada segmento corporal. O estudo científico dos diplópodes designa-se como diplopodologia.
Índice
1 Etimologia e nomes
2 Taxonomia
2.1 Classificação
2.2 Evolução
2.3 Grupos vivos
2.4 Registo fóssil
2.5 Relação com outros miriápodes
2.6 Distinção com os centípedes
3 Características
3.1 Cabeça
3.2 Corpo
3.3 Órgãos internos
4 Reprodução e crescimento
5 Ecologia
5.1 Habitat e distribuição
5.2 Hábitos de escavação
5.3 Dieta
5.4 Predadores e parasitas
5.5 Mecanismos de defesa
5.6 Outras interacções interespecíficas
6 Interacção com as pessoas
7 Referências
8 Ver também
9 Ligações externas
Etimologia e nomes |
O nome "Diplopoda" deriva do grego antigo διπλοῦς diplous, 'duplo' e ποδός, podos, 'pés', referindo-se ao facto de apresentarem dois pares de patas na maioria dos segmentos. O nome comum que se lhes dá em muitos idiomas de "milípedes" (mil pés) faz referência à sua grande quantidade de patas (superior à dos centípedes, "cem pés"), embora nenhum chegue a mil.[5][6]
Taxonomia |
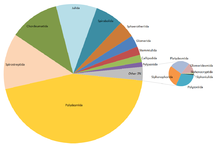
Diversidade relativa aproximada das ordens de milípedes existentes, que varia desde cerca de 3 500 espécies de Polydesmida a 2 espécies de Siphoniulida.[7]
Foram descritas aproximadamente 12 000 espécies de diplópodes. A estimativa do número total de espécies que poderia haver ao todo é de entre 15 000[8] e 80 000.[9] Algumas das espécies estão amplamente distribuídas; têm mais capacidades de dispersão, ao dependerem da locomoção terrestre e dos habitats húmidos. Estes factores favoreceram o isolamento genético e a rápida especiação, produzindo muitas linhagens com áreas de distribuição restritas.[10]
Os membros vivos dos diplópodes dividem-se em 16 ordens em duas subclasses.[7] A subclasse basal Penicillata compreende uma só ordem, Polyxenida. Os demais milípedes pertencem à subclasse Chilognatha, que consta de duas infraclasses: Pentazonia, que contém os Oniscomorpha, e os Helminthomorpha, que incluem a maioria das espécies.[11][12]
Classificação |
A classificação de alto nível dos diplópodes é apresentada mais abaixo e baseia-se em Shear, 2011,[7] e Shear & Edgecombe, 2010[13] (grupos extintos). Recentes estudos cladísticos e moleculares mudaram os esquemas da classificação tradicional indicados antes, e em particular a posição das ordens Siphoniulida e Polyzoniida não está ainda estabelecida.[9] A situação e posições dos grupos extintos (†), conhecidos apenas pelos seus fósseis, são provisórias e não estão completamente resolvidas.[9][13] Depois de cada nome indica-se a autoridade (nome da pessoa que lhe deu nome ou definiu originalmente).
Classe Diplopoda de Blainville in Gervais, 1844
- Subclasse Penicillata Latrielle, 1831
- Ordem Polyxenida Verhoeff, 1934
- Ordem Polyxenida Verhoeff, 1934
- Subclasse †Arthropleuridea (situado en Penicillata por alguns autores)[13]
- Ordem †Arthropleura Waterlot, 1934
- Ordem †Eoarthropleura Shear & Selden, 1995
- Ordem †Microdecemplicida Wilson & Shear, 2000
- Ordem †Arthropleura Waterlot, 1934
- Subclasse Chilognatha Latrielle, 1802
- Ordem †Zosterogrammida Wilson, 2005 (Chilognatha incertae sedis)[13]
- Infraclasse Pentazonia Brandt, 1833
- Ordem †Amynilyspedida Hoffman, 1969
- Superordem Limacomorpha Pocock, 1894
- Ordem Glomeridesmida Cook, 1895
- Ordem Glomeridesmida Cook, 1895
- Superordem Oniscomorpha Pocock, 1887
- Ordem Glomerida Brandt, 1833
- Ordem Sphaerotheriida Brandt, 1833
- Ordem Glomerida Brandt, 1833
- Ordem †Amynilyspedida Hoffman, 1969
- Infraclasse Helminthomorpha Pocock, 1887
- Superordem †Archipolypoda Scudder, 1882
- Ordem †Archidesmida Wilson & Anderson 2004
- Ordem †Cowiedesmida Wilson & Anderson 2004
- Ordem †Euphoberiida Hoffman, 1969
- Ordem †Palaeosomatida Hannibal & Krzeminski, 2005
- Ordem †Archidesmida Wilson & Anderson 2004
- Ordem †Pleurojulida Schneider & Werneburg, 1998 (possivelmente irmã de Colobognatha)[9]
- Subterclasse Colobognatha Brandt, 1834
- Ordem Platydesmida Cook, 1895
- Ordem Polyzoniida Cook, 1895
- Ordem Siphonocryptida Cook, 1895
- Ordem Siphonophorida Newport, 1844
- Ordem Platydesmida Cook, 1895
- Subterclasse Eugnatha Attems, 1898
- Superordem Juliformia Attems, 1926
- Ordem Julida Brandt, 1833
- Ordem Spirobolida Cook, 1895
- Ordem Spirostreptida Brandt, 1833
- Superfamília †Xyloiuloidea Cook, 1895 (às vezes alinhada com Spirobolida)[14]
- Ordem Julida Brandt, 1833
- Superordem Nematophora Verhoeff, 1913
- Ordem Callipodida Pocock, 1894
- Ordem Chordeumatida Pocock 1894
- Ordem Stemmiulida Cook, 1895
- Ordem Siphoniulida Cook, 1895
- Ordem Callipodida Pocock, 1894
- Superordem Merocheta Cook, 1895
- Ordem Polydesmida Pocock, 1887
- Ordem Polydesmida Pocock, 1887
- Superordem Juliformia Attems, 1926
- Superordem †Archipolypoda Scudder, 1882
- Ordem †Zosterogrammida Wilson, 2005 (Chilognatha incertae sedis)[13]
Evolução |
Os milípedes estão entre os primeiros animais que colonizaram a Terra durante o período Silúrico.[15] As formas iniciais provavelmente comiam musgos e plantas vasculares primitivas. Há dois grupos maiores de milípedes cujos membros estão todos extintos: os Archipolypoda, que incluem os animais terrestres mais antigos conhecidos, e os Arthropleuridea, que contêm os invertebrados terrestres mais compridos conhecidos. A criatura terrestre mais antiga da qual temos conhecimento é a Pneumodesmus newmani, que media 1 cm de comprimento e era um Archipolypoda, que viveu há 428 milhões de anos no Silúrico superior e que apresenta claras evidências de espiráculos (orifícios respiratórios), o que testemunha os seus hábitos respiratórios.[13][16] Durante o Carbonífero superior (entre 340 e 280 milhões de anos atrás), Arthropleura converteu-se no maior invertebrado da vida terrestre de que se tenha registo, alcançando um comprimento de pelo menos 2 m.[17] Os milípedes também mostram a primeira evidência de defesa química, uma vez que alguns fósseis devónicos têm aberturas de glândulas defensivas chamadas ozóporos.[13] Os milípedes, centípedes e outros artrópodes terrestres antigos atingiram grandes dimensões comparativamente com as modernas espécies, graças aos ambientes ricos em oxigénio do Devónico e Carbonífero, e alguns inclusivamente podiam ultrapassar um metro de comprimento. À medida que os níveis de oxigénio se reduziram com o tempo, os artrópodes tornaram-se mais pequenos.[18]
Grupos vivos |

Octoglena sierra (Colobognatha, Polyzoniida)

Anadenobolus monilicornis (Juliformia, Spirobolida)

Harpaphe haydeniana (Polydesmida)
A história da classificação científica dos milípedes começou com Carl Linnaeus, que na sua 10ª edição do Systema Naturae, 1758, nomeou sete espécies de Julus como "Insecta Aptera" (insectos sem asas).[19] Em 1802, o zoólogo francês Pierre André Latreille propôs o nome de Chilognatha para o primeiro grupo, que agora se chama Diplopoda, e em 1840 o naturalista alemão Johann Friedrich von Brandt produziu as primeiras classificações detalhadas. O próprio nome Diplopoda foi cunhado em 1844 pelo zoólogo francês Henri Marie Ducrotay de Blainville. Entre 1890 e 1940, a taxonomia dos milípedes foi impulsionada por um número relativamente reduzido de investigadores, e os maiores contributos foram realizados por Carl Attems, Karl Wilhelm Verhoeff e Ralph Vary Chamberlin, cada um dos quais descreveu cerca de 1 000 espécies, e por Orator F. Cook, Filippo Silvestri, R. I. Pocock e Henry W. Brölemann.[9] Este foi um período no qual a ciência da diplopodologia floresceu: os números de descrições de espécies foram, em média, os mais altos da história, por vezes excedendo as 300 por ano.[8]
Em 1971, o biólogo holandês C. A. W. Jeekel publicou uma lista completa de todos os géneros e famílias dos milípedes descritos entre 1758 e 1957 no seu Nomenclator Generum et Familiarum Diplopodorum, um trabalho que se considera ter dado começo à "era moderna" da taxonomia dos milípedes.[20][21] Em 1980, o biólogo americano Richard L. Hoffman publicou uma classificação dos milípedes que reconhecia já os Penicillata, Pentazonia e Helminthomorpha,[22] e a primeira análise filogenética das ordens de milípedes utilizando métodos cladísticos modernos foi publicada em 1984 por Henrik Enghoff na Dinamarca.[23] Uma classificação de 2003 feita pelo americano Rowland Shelley assemelha-se em muito à proposta feita originalmente por Verhoeff, e continua sendo o esquema de classificação actualmente aceito (mostrado abaixo), apesar dos estudos moleculares mais recentes que propõem relações que entram em conflito com estas.[9][13] Um resumo de 2011 da diversidade de famílias dos milípedes de William A. Shear situou a ordem Siphoniulida dentro do grande grupo dos Nematomorpha.[7]
| Diplopoda |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Registo fóssil |
Para além das 16 ordens vivas, existem 9 ordens extintas e uma superfamília conhecida apenas a partir de fósseis. As relações entre estes grupos vivos são controversas. Os extintos Arthropleuridea foram durante um longo período considerados como uma classe de miriápodes própria, embora os trabalhos feitos nos inícios do século XXI estabelecessem que o grupo se tratava duma subclasse de milípedes.[24][25][26] Várias ordens vivas aparecem também no registo fóssil. Abaixo são apresentadas duas classificações dos grupos de milípedes fósseis.[9][13] Os grupos extintos estão acompanhados pelo símbolo †. Não é mostrada a ordem extinta dos Zosterogrammidae, alguns quilógnatos de posição incerta na classificação.[13]
| |||||||||||||||||||||||||||
| Hipótese alternativa sobre as relação dos fósseis.[9][25] |
| Diplopoda |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Relação com outros miriápodes |

Acredita-se que os paurópodes sejam os parentes mais próximos dos milípedes
Embora as relações entre as ordens dos milípedes ainda sejam motivo de debate, a classe Diplopoda no conjunto é considerada um grupo monofilético de artrópodes: todos os milípedes estão mais relacionados entre si do que com qualquer outro artrópode. Diplopoda é uma classe dentro do subfilo Myriapoda (miriápodes) dos artrópodes, que incluem os centípedes (classe Chilopoda) e os menos conhecidos paurópodes (classe Pauropoda) e sínfilos (classe Symphyla). Dentro dos miriápodes, considera-se desde há algum tempo que os parentes mais próximos ou grupo irmão dos milípedes sejam os paurópodes, os quais possuem um collum e diplossegmentos.[9]
Distinção com os centípedes |
Estabelecer as diferenças entre os milípedes e centípedes é uma questão básica para distinguir os grupos de miriápodes.[27] Os dois grupos de miriápodes compartilham algumas parecenças, como ter corpos multissegmentados, muitas patas, um só par de antenas e a posse de órgãos de Tömösváry, mas têm muitas diferenças e diferentes histórias evolutivas, uma vez que o antepassado comum mais recente dos centípedes e milípedes viveu por volta de 450 a 475 milhões de anos atrás, no período Silúrico.[28] A cabeça por si só exemplifica as diferenças: os milípedes têm antenas curtas, dobradas formando um ângulo ou cotovelo para examinar o substrato, e um par de robustas mandíbulas, um único par de maxilas fundidas formando um lábio; já os centípedes possuem antenas compridas sem formação de cotovelo, um par de mandíbulas, dois pares de maxilas e um par de garras grandes e venenosas.[29][30]

Representantes característicos dos milípedes e centípedes (não necessariamente à escala)
| Característica | Milípedes | Centípedes |
|---|---|---|
| Patas | Dois pares na maioria dos segmentos; unidas à parte lateral do corpo | Um par por segmento corporal; unidas às laterais do corpo; o último par estende-se para trás |
| Locomoção | Geralmente adaptados para enterrarem-se ou habitar em pequenas buracos; movimentos lentos | Geralmente adaptados para correr, excepto os Geophilomorpha (centípedes do solo), que se enterram |
| Alimentação | Principalmente detritívoros, alguns herbívoros, poucos carnivoros; sem veneno | Principalmente carnivoros com garras modificadas como se fossem caninos e com veneno |
| Espiráculos | Na parte inferior do corpo | Nas laterais ou na parte superior do corpo |
| Orifícios reprodutores | Terceiro segmento corporal | Último segmento corporal |
| Comportamento reprodutor | Os machos geralmente inserem espermatóforos nas fêmeas com gonóporos | Os machos produzem espermatóforos que normalmente são recolhidos pelas fêmeas |
Características |

Tipos corporais representativos dos Penicillata (acima), Pentazonia (no meio) e Helminthomorpha (abaixo)

Anatomia anterior de um milípede helmintomorfo generalizado

Detalhe da cabeça dum diplópode, em Santiago de Compostela
Os milípedes têm diversas formas e tamanhos corporais, que variam de 2 mm até cerca 35 cm de comprimento,[31] e podem ter entre 11 a 100 segmentos corporais.[32] Geralmente são de cor preta ou castanha, embora existam algumas espécies com cores berrantes, e algumas até com cores aposemáticas como aviso de que são tóxicas.[6] As espécies de Motyxia produzem cianeto como defesa química e são bioluminescentes.[33]
Os estilos corporais variam muito entre os principais grupos de milípedes. Na subclasse basal dos pequenos Penicillata o exoesqueleto é mole e não calcificado, e está coberto de pelos ou sedas proeminentes. Os demais milípedes pertencem à subclasse dos Chilognatha, e têm exoesqueletos endurecidos. Os quilógnatos por sua vez dividem-se em duas infraclasses: os Pentazonia possuem grupos de corpos relativamente curtos como os Oniscomorpha e os Helminthomorpha, que englobam a grande maioria das espécies, têm corpos compridos com muitos segmentos.[11][12]
Cabeça |
A cabeça dos milípedes é tipicamente arredondada em cima e aplanada embaixo e é constituída por um par de grandes mandíbulas na frente duma estrutura semelhante a uma placa chamada gnatoquilário ("lábio da mandíbula").[9] A cabeça contém um só par de antenas com sete ou oito segmentos e um grupo de cones sensoriais na extremidade. Muitas ordens também possuem um par de órgãos sensoriais chamados órgãos de Tömösváry, com forma de anéis ovais na parte posterior e lateral à base das antenas. Desconhece-se quais as suas funções,[9] mas também aparecem em alguns centípedes, e são utilizados possivelmente para medir os níveis de humidade ou luz no ambiente que os rodeia.[34]
Os olhos dos milípedes consistem de vários ocelos com lentes planas simples dispostas num grupo ou mancha em cada lado da cabeça. Estas manchas também se chamam campos oculares ou ocelários. Muitas espécies de milípedes, incluindo toda a ordem dos Polydesmida e milípedes que habitam tocas como os Causeyella e Trichopetalum, teriam antepassados que poderiam ver mas que perderam mais tarde os olhos, tornando-se cegos.[31]
Corpo |

Comparação (sem escala) das expansões laterais do noto (paranoto) em polidésmidos (esquerda) e platidésmidos (direita)
Os corpos dos diplópodes são formados por dois tagmas, cabeça e tronco, estrutura básica dos miriápodes. Os corpos podem ser aplanados ou cilíndricos e são compostos por numerosos segmentos metaméricos, cada um com um exoesqueleto formado por cinco placas quitinosas, que são: uma única placa dorsal (o tergito), uma em cada lado (pleuritos), e uma na parte ventral (esternito) onde se unem as patas. Em muitos diplópodes, estas placas estão fundidas em vários níveis, formando por vezes um único anel cilíndrico. Normalmente as placas são duras, uma vez que estão impregnadas de sais cálcicos.[32] Dado que carecem de uma cutícula cerosa, estão susceptíveis à perda de água e precisam passar a maior parte do seu tempo em ambientes húmidos.[35]
O primeiro segmento por trás da cabeça não tem patas e denomina-se colum (colo ou pescoço).[16] O segundo, terceiro e quarto segmentos do corpo comportam um único par de patas e denominam-se "haplossegmentos" (estes três haplossegmentos são por vezes chamados de "tórax"). Os segmentos restantes, do quinto ao posterior, denominam-se diplossegmentos ou segmentos duplos, formados pela fusão de dois segmentos embrionais. Cada diplossegmento comporta dois pares de patas. Nalguns diplópodes, os últimos segmentos podem não ter patas. Os termos "segmento" ou "anel corporal" utilizam-se amiúdo indistintamente para referir-se tanto a haplossegmentos como a diplossegmentos. O último segmento denomina-se télson e consiste num anel pré-anal sem patas, um par de válvulas anais (placas que se podem fechar ao redor do ânus), e uma pequena escama debaixo do ânus.[9][32]
Os diplópodes de várias ordens têm extensões em forma de quilha da parede corporal conhecidas como paranotos, que podem variar amplamente em forma, tamanho e textura; as modificações incluem lobos, papilas, cristas, espinhas e amossegas.[6] Os paranotos podem permitir aos diplópodes incrustar-se com maior segurança nas fendas, proteger as patas ou fazer com que sejam difíceis de serem devorados pelos predadores.[36]
As patas são constituídas por sete segmentos e unidas à parte inferior do corpo. As patas dum só indivíduo são geralmente bastante parecidas umas às outras, embora costumem ser mais compridas nos machos do que nas fêmeas, e os machos dalgumas espécies podem ter um primeiro par de patas reduzido ou alongado.[37] As mais mudanças de maior destaque das patas têm que ver com a reprodução, conforme falaremos mais adiante (transformação em gonóporos). Apesar do seu nome comum de milípedes, não se conhece nenhum milípede que tenha mil patas: as espécies comuns têm entre 34 e 400 patas e o maior de que se tem registo é o da espécie Illacme plenipes, com alguns indivíduos com cerca de 750 patas, o que é mais do que qualquer outra criatura da Terra.[38]

Fêmea de Illacme plenipes com 618 patas (309 pares)
Órgãos internos |
Os milípedes respiram por dois pares de espiráculos localizados ventralmente em cada segmento perto da base das patas.[27] Cada abertura concede passagem a uma cavidade interna que pode armazenar oxigénio e conecta com um sistema traqueal.[39] O coração estende-se por todo o comprimento do corpo, com uma aorta que se estende até à cabeça. Os órgãos excretores são dois pares de túbulos malpighianos, localizados perto da parte do meio do tracto digestivo. O tracto digestivo consiste num simples tubo com dois pares de glándulas salivares que ajudam a digerir a comida.[32]
Reprodução e crescimento |
Os diplópodes apresentam diversos estilos de acasalamento e estruturas. Na ordem basal dos Polyxenida o acasalamento é indirecto: os machos depositam espermatóforos em redes que segregam com algumas glándulas especiais, e os espermatóforos são depois recolhidos pelas fêmeas. Noutros grupos de diplópodes, os machos possuem um ou dois pares de patas modificadas chamadas gonóporos, que utilizam para transferir o esperma à fêmea durante a cópula. A localização dos gonóporos difere entre os grupos: nos machos de Pentazonia situam-se na parte posterior do corpo e denominam-se telópodes, e podem também servir para agarrar as fêmeas, enquanto que nos Helminthomorpha (que compreendem a grande maioria das espécies) estão situados no sétimo segmento corporal.[9] Algumas espécies são partenogenéticas, e possuem poucos ou nenhum macho.[40]
@media all and (max-width:720px){.mw-parser-output .tmulti>.thumbinner{width:100%!important;max-width:none!important}.mw-parser-output .tmulti .tsingle{float:none!important;max-width:none!important;width:100%!important;text-align:center}}


Os gonóporos podem ter diferentes formas e tamanhos, e em aspecto podem variar tanto que as patas locomotoras, por se tratarem de estruturas tão complexas, acabam por ser completamente diferentes das patas a que estamos habituados. Em alguns grupos, os gonóporos mantêm-se retraídos dentro do corpo; noutros projectam-se para a frente em paralelo ao corpo. A morfologia dos gonóporos é a forma predominante de determinar a espécie entre os milípedes: as estruturas podem divergir grandemente entre espécies que estão intimamente relacionadas, por outro lado podem divergir muito pouco dentro duma única espécie.[41] Os gonóporos desenvolvem-se gradualmente a partir de patas locomotoras ao longo das sucessivas mudas (ecdises) até à maturação reprodutora.[42]
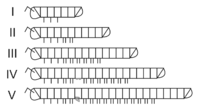
Estádios do crescimento de Nemasoma (Nemasomatidae), que atinge a capacidade reprodutiva no estádio V
Os orifícios genitais (gonóporos) de ambos os sexos estão localizados na parte lateral do terceiro segmento corporal (perto do segundo par de patas) e podem estar acompanhados no macho por um ou dois pénis, que depositam os pacotes de esperma nos gonóporos. Na fêmea, os poros genitais abrem-se em pequenos sacos pares chamados cifópodes ou vulvas, que estão cobertos por pequenas tampas em forma de capacete, e são utilizados para armazenar o esperma após a cópula.[32] A morfologia dos cifópodes pode também ser utilizada para identificar as espécies. Os espermatozoides dos milípedes carecem de flagelos, uma característica exclusiva destes animais dentro dos miriápodes.[9]
Em todos excepto nos Polyxenida, a copulação ocorre quando os dois indivíduos se põem um de frente para o outro. A copulação pode ser precedida por determinados comportamentos do macho, como golpear com as antenas, correr ao longo do dorso da fêmea, oferecer secreções glandulares comestíveis, ou, no caso dos Oniscomorpha, a estridulação.[43] Durante a cópula na maioria dos diplópodes, o macho situa o seu sétimo segmento em frente do terceiro segmento da fêmea, e insere os seus gonóporos para fazer sair as vulvas, antes de dobrar o seu corpo para depositar o esperma nos seus gonóporos e reinserir os gonóporos "carregados" no corpo da fêmea.[37]
As fêmeas põem entre 10 a 300 ovos de cada vez, dependendo da espécie, fertilizando-os, no momento em que os põem, com o esperma que armazenaram. Muitas espécies depositam os ovos sobre o solo húmido ou detritos orgânicos, mas outras constroem ninhos forrados com fezes secas, e podem proteger os ovos em casulos de seda.[32] Na maioria das espécies, a fêmea abandona os ovos depois de os pôr, mas algumas espécies das ordens Platydesmida e Stemmiulida cuidam dos ovos e das suas ninhadas.[27]
As ninhadas eclodem em poucas semanas, e têm tão-somente três pares de patas, seguidas de até quatro segmentos sem patas. À medida que crescem, mudam sucessivamente, acrescentando mais segmentos e patas em cada muda. Algumas espécies mudam em câmaras especialmente preparadas, no solo ou de seda,[44] e podem também refugiar-se nelas durante o tempo chuvoso, e a maioria das espécies come o exoesqueleto velho descartado após a muda. A fase adulta, quando os indivíduos atingem a capacidade reprodutiva, é geralmente alcançada no estádio da muda final, o qual varia entre espécies e ordens, embora algumas espécies continuem as mudas depois de se tornarem adultas. Para além disso, algumas espécies alternam entre estádios reprodutores e não reprodutores depois da maturação, um fenómeno chamado periodomorfose, no qual as estruturas reprodutoras sofrem regressão durante os estádios não reprodutores.[40] Os milípedes podem viver entre um e dez anos, dependendo da espécie.[32]
Ecologia |
Habitat e distribuição |
Os milípedes vivem em todos os continentes excepto na Antártida e ocupam quase todos os habitats terrestres, desde o Círculo Polar Ártico na Islândia, Noruega e Rússia, até ao sul na Província de Santa Cruz, Argentina.[45][46] Normalmente habitam os solos florestais, nos quais vivem sob as camadas de folhas caídas, em madeira podre ou no solo, com predilecção por condições húmidas. É um animal bastante comum nos pátios de compostagem de resíduos orgânicos, sendo um dos organismos responsáveis pela transformação desses materiais em adubo orgânico.[47] Em zonas moderadas, os milípedes são mais abundantes nas florestas caducas húmidas, e podem atingir densidades de 1 000 indivíduos por metro quadrado. Outros habitats são as florestas de coníferas, desertos, covas e ecossistemas alpinos.[27][46] Algumas espécies podem sobreviver às inundações de água doce e viver submersos debaixo de água por cerca de 11 meses.[48][49] Algumas espécies vivem perto da costa marítima e podem sobreviver em condições parcialmente salinas.[40][50]
Hábitos de escavação |
Os diplossegmentos dos milípedes evoluíram em conjunção com os seus hábitos de escavação, e quase todos os milípedes adoptam um estilo de vida essencialmente subterrâneo. Usam três métodos principais de soterramento: escavar, acunhar-se e perfurar. Os das ordens Julida, Spirobolida e Spirostreptida baixam as cabeças e metem-se à força no seu substrato, e o collum é a porção do seu exoesqueleto que colocam primeiro. Os milípedes de dorso aplanado da ordem Polydesmida tendem a inserir o seu extremo dianteiro, como uma cunha, numa fenda horizontal, e depois ensancham a fenda empurrando para a frente com as patas; os seus paranotos neste exemplo constituem a principal superfície de elevação. A técnica de perfuração é utilizada pelos membros da ordem Polyzoniida, que têm segmentos menores na parte dianteira e cada vez mais compridos na extremidade traseira; propulsionam-se para diante nas fendas com as patas, e o corpo com forma de cunha ensancha o oco à medida que avançam. Alguns milípedes adoptaram um estilo de vida sobre a superfície do chão, pelo que perderam os seus hábitos de soterrar-se. Isto pode dever-se ao facto de que são demasiado pequenos para fazer força suficiente para escavar e soterrar-se ou porque são demasiado grandes para fazer o esforço valer-lhes a pena, ou nalguns casos porque se movem relativamente rápido (para um milípede) e são predadores activos.[6]
Dieta |
A maioria dos milípedes são detritívoros e alimentam-se de vegetação em decomposição, fezes ou matéria orgânica misturada com o solo. Desempenham um importante papel na degradação e decomposição das folhas caídas: estima-se que a taxa de consumo duma espécie qualquer varie de 1 a 11% de todas as folhas caídas, dependendo da espécie e da região, e em conjunto os milípedes podem consumir quase todas as folhas caídas duma região. As folhas caídas são fragmentadas no tubo digestivo dos milípedes e excretadas na forma de bolinhas de fragmentos de folhas, algas, fungos e bactérias, o que facilita a decomposição pelos micro-organismos.[37] Nos chãos das florestas tropicais onde as populações de minhocas são baixas, os milípedes desempenham um importante papel ao facilitarem a decomposição microbiana das folhas.[6] Alguns milípedes são herbívoros, alimentando-se de plantas vivas, e certas espécies podem ser graves pragas para a agricultura nalguns países. Os milípedes da ordem Polyxenida comem algas das crostas das árvores e as Platydesmida alimentam-se de fungos.[9] Algumas espécies são omnívoras ou ocasionalmente carnívoras, alimentando-se de insectos, centípedes, minhocas ou caracóis.[32][51] Algumas espécies têm peças bucais perfuradoras com as quais podem sugar o sumo das plantas.[27]
Predadores e parasitas |
Os milípedes são predados por uma ampla variedade de animais, incluindo vários répteis, anfíbios, aves, mamíferos e insectos.[9] Os predadores mamíferos como os quatis e suricatas fazem rolá-los pelo chão e capturam milípedes para que libertem as suas secreções defensivas e esfregam as ditas secreções antes de comê-los;[52] acredita-se que algumas rãs dendrobátidas incorporam os compostos tóxicos dos milípedes para a sua própria defesa.[53] Vários invertebrados têm comportamentos especializados ou estruturas para alimentar-se de milípedes, entre eles larvas de escaravelhos Phengodidae,[54] as formigas Probolomyrmex,[55] as lesmas Chlamydephorus[56] e os escaravelhos Sceliages e Deltochilum.[57][58] Uma grande subfamília de Reduviidae, a Ectrichodiinae, com cerca de 600 espécies, especializou-se na predação de milípedes.[59] Entre os parasitas dos milípedes existem os nematódeos, moscas feomíidas e acantocéfalos.[9]
Mecanismos de defesa |

Ammodesmus nimba da Guiné, África ocidental, enroscado em posição defensiva
Por não terem grande velocidade e pela sua incapacidade de morder ou picar, o principal mecanismo de defesa dos milípedes é enroscar-se formando uma espiral apertada, projectando o mais possível as suas delicadas patas para dentro do seu exoesqueleto encouraçado.[60]
Muitas espécies também emitem como defesa secundária várias secreções líquidas de mau cheiro através de poros microscópicos chamados ozóporos (que são as aberturas das suas glándulas "odoríferas" ou "repugnatórias"), situados nas partes laterais dos seus corpos. Entre os muitos compostos químicos irritantes ou tóxicos que se encontraram nestas secreções estão alcaloides, benzoquinonas, fenóis, terpenoides e cianeto de hidrogénio.[61][62] Algumas destas substâncias são cáusticas e podem queimar o exoesqueleto das formigas e outros insectos predadores, e a pele e olhos de predadores de maior porte. Podem ser vistos primatas como os macacos-pregos (Cebinae) e lémures a irritar intencionalmente os milípedes para esfregar as suas secreções químicas sobre o seu corpo com a intenção de repelir os mosquitos.[63][64][65] Alguns destes compostos defensivos também apresentam uma actividade antifúngica.[66]
Os milípedes da ordem Polyxenida não possuem exoesqueleto encouraçado nem glándulas odoríferas, estando, por sua vez, cobertos por numerosos pelos ou sedas que pelo menos na espécie Polyxenus fasciculatus podem desprender-se e enredar as formigas.[67]
Outras interacções interespecíficas |
Alguns milípedes estabelecem relações mutualistas com organismos doutras espécies, nas quais ambas as espécies beneficiam da interacção, ou relações de comensalismo, nas quais uma única espécie beneficia, enquanto que a outra não se vê afectada. Várias espécies formam estreitas relações com as formigas, uma relação chamada mirmecofilia, especialmente na família Pyrgodesmidae (Polydesmida), que contém "mirmecófilos obrigados", espécies que só se encontraram em colónias de formigas. A maioria das espécies são "mirmecófilos facultativos", uma vez que não estão exclusivamente associadas com formigas, incluindo muitas espécies de Polyxenida, que se encontraram em formigueiros de todo o mundo.[68]
Muitas espécies de milípedes apresentam relação de comensalismo com ácaros das ordens Mesostigmata e Astigmata. Acredita-se que muitos destes ácaros sejam foréticos em vez de parasitas, o que significa que utilizam os hóspedes milípedes para a sua dispersão.[69][70]
Em 2011 foi descrita uma nova interacção entre milípedes e musgos, na qual os indivíduos da nova espécie descoberta Psammodesmus bryophorus têm dez espécies de musgos que vivem na sua superfície dorsal, graças a que o milípede fica camuflado e potencia a dispersão dos musgos.[71][72]
Interacção com as pessoas |
Os milípedes geralmente apresentam pouco impacto na economia humana ou no bem-estar das pessoas, principalmente quando em comparação com os insectos, mas localmente podem se tornar numa moléstia ou numa praga agrícola. Os milípedes não mordem nem picam, e as suas secreções defensivas são quase sempre inofensivas para os humanos (geralmente causam só descolorações menores na pele), embora as secreções de algumas espécies tropicais possam causar dor, prurido, eritema local, edema, ampolas, eczema e, ocasionalmente, pele gretada.[73][74][75][76] Se as excreções atingem os olhos causam uma irritação geral e efeitos potencialmente mais graves como conjuntivite e queratite.[77]

Blaniulus guttulatus pode ser uma praga agrícola
Alguns centípedes são considerados pragas domésticas nalguns países, incluindo o Xenobolus carnifex, que pode infestar os telhados de palha na Índia,[78] e o Ommatoiulus moreleti, que periodicamente invade as casas na Austrália. Outras espécies mostram um comportamento gregário (enxameiam-se) periodicamente, o que pode provocar invasões domésticas,[79] danos agrícolas[80] ou mesmo atrasos nos transportes ferroviários sempre que as vias ficam escorregadias por causa dos restos esmagados de centenas de milípedes.[37][81][82] Alguns milípedes podem prejudicar significativamente as colheitas: o Blaniulus guttulatus é uma praga para as colheitas de beterraba-sacarina e outras plantas das culturas sachadas..[40]
Alguns dos maiores milípedes da ordem Spirobolida, Spirostreptida e Sphaerotheriida tornaram-se mascotes populares em certos países.[83] Algumas das espécies que se vendem como mascotes são as dos géneros Archispirostreptus, Aphistogoniulus, Narceus e Orthoporus.[84]
Os milípedes aparecem no folclore e medicina tradicional de todo o mundo. Algumas culturas associam a actividade dos milípedes com a chegada das chuvas.[85] Na cultura iorubá da Nigéria, os milípedes são utilizados em rituais relacionados com a gravidez e os negócios, e os milípedes quando esmagados são utilizados para tratamento da febre, unheiros e convulsões em crianças.[86] Na Zâmbia, a polpa dos milípedes esmagados é utilizada para tratar ferimentos, e o povo Bafia dos Camarões utiliza o sumo de milípedes para tratar as dores de ouvidos.[85] Em determinadas tribos Bhotiya do Himalaia o fumo da queima de milípedes secos utiliza-se para tratar as hemorroidas.[87] Os povos nativos da Malásia usam as secreções dos milípedes para envenenamento de setas.[85] Observou-se que as secreções do Spirobolus bungii inibem a divisão de células cancerígenas humanas.[88] O único uso registado de milípedes como alimento humano deu-se no povo Bobo de Burquina Faso, que come milípedes secos cozidos com molho de tomate.[89]
Os milípedes também servem de inspiração para algumas investigações científicas. Em 1963, desenhou-se um veículo que caminhava em 36 patas, feito com base na locomoção dos milípedes.[90] Alguns robôs experimentais tiveram a mesma inspiração,[91][92] em especial quando são necessários para transportar cargas pesadas por terrenos difíceis e com curvas.[93] Em biologia, alguns autores advogam pelo uso de milípedes como organismos-modelo para o estudo da fisiologia dos artrópodes e os processos de desenvolvimento que controlam o número e forma dos segmentos corporais.[37]
Referências
↑ «Diplópode». Priberam. Consultado em 14 de Julho de 2018
↑ «Piolho-de-cobra: conheça curiosidades sobre este animal». Natureza e Conservação. Consultado em 10 de agosto de 2017
↑ ab «Gongolo». Michaelis On-Line. Consultado em 7 de agosto de 2017
↑ Brewer, Michael S.; Sierwald, Petra; Bond, Jason E. (15 de maio de 2012). «Millipede Taxonomy after 250 Years: Classification and Taxonomic Practices in a Mega-Diverse yet Understudied Arthropod Group». PLoS ONE. 7 (5). ISSN 1932-6203. PMID 22615951. doi:10.1371/journal.pone.0037240
↑ Hoffman, Richard L. (1990). «Diplopoda». In: Dindal, Daniel L. Soil Biology Guide. [S.l.]: John Wiley & Sons. p. 835. ISBN 978-0-471-04551-9
↑ abcde Ruppert, Edward E.; Fox, Richard, S.; Barnes, Robert D. (2004). Invertebrate Zoology, 7th edition. [S.l.]: Cengage Learning. pp. 711–717. ISBN 978-81-315-0104-7
↑ abcd Shear, W. (2011). «Class Diplopoda de Blainville in Gervais, 1844. In: Zhang, Z.-Q. (Ed.) Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness» (PDF). Zootaxa. 3148: 159–164
↑ ab Brewer, Michael S.; Sierwald, Petra; Bond, Jason E. (2012). «Millipede taxonomy after 250 years: Classification and taxonomic practices in a mega-diverse yet understudied arthropod group». PLoS ONE. 7 (5): e37240. Bibcode:2012PLoSO...737240B. PMC 3352885 . PMID 22615951. doi:10.1371/journal.pone.0037240
. PMID 22615951. doi:10.1371/journal.pone.0037240
↑ abcdefghijklmnopq Sierwald, Petra; Bond, Jason E. (2007). «Current status of the myriapod class Diplopoda (Millipedes): Taxonomic diversity and phylogeny». Annual Review of Entomology. 52 (1): 401–420. PMID 17163800. doi:10.1146/annurev.ento.52.111805.090210
↑ Barker, G.M. (2004). Natural Enemies of Terrestrial Molluscs. [S.l.]: CABI. pp. 405–406. ISBN 978-0-85199-061-3
↑ ab Bueno-Villegas, Julián; Sierwald, Petra; Bond, Jason E. «Diplopoda». In: Bousquets, J. L.; Morrone, J. J. Biodiversidad, taxonomia y biogeografia de artropodos de Mexico (PDF). [S.l.: s.n.] pp. 569–599
↑ ab Shelley, Rowland M. «Millipedes». University of Tennessee: Entomology and Plant Pathology. Consultado em 17 de julho de 2016
↑ abcdefghi Shear, William A.; Edgecombe, Gregory D. (2010). «The geological record and phylogeny of the Myriapoda». Arthropod Structure & Development. 39 (2–3): 174–190. PMID 19944188. doi:10.1016/j.asd.2009.11.002
↑ Hoffman, R. L. (1963). «New genera and species of Upper Paleozoic Diplopoda». Journal of Paleontology. 37 (1): 167–174. JSTOR 1301419
↑ Garwood, Russell; Edgecombe, Gregory (2011). «Early terrestrial animals, evolution and uncertainty». Evolution, Education, and Outreach. 4 (3): 489–501. doi:10.1007/s12052-011-0357-y
↑ ab Wilson, Heather M.; Anderson, Lyall I. (2004). «Morphology and taxonomy of Paleozoic millipedes (Diplopoda: Chilognatha: Archipolypoda) from Scotland». Journal of Paleontology. 78 (1): 169–184. doi:10.1666/0022-3360(2004)078<0169:MATOPM>2.0.CO;2
↑ Sues, Hans-Dieter (15 de janeiro de 2011). «Largest Land-Dwelling "Bug" of All Time». National Geographic. Consultado em 25 de fevereiro de 2016. Arquivado do original em 4 de março de 2016
↑ Lockley, M. G.; Meyer, Christian (2013). «The Tradition of Tracking Dinosaurs in Europe». Dinosaur Tracks and Other Fossil Footprints of Europe. [S.l.]: Columbia University Press. pp. 25–52. ISBN 978-0-231-50460-7
↑ Caroli Linnaei (1758). Systema naturae per regna tria naturae: secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. [S.l.: s.n.] pp. 639–640
↑ Shelley, R. M. (2007). «Taxonomy of extant Diplopoda (Millipeds) in the modern era: Perspectives for future advancements and observations on the global diplopod community (Arthropoda: Diplopoda)» (PDF). Zootaxa. 1668: 343–362
↑ Shelley, Rowland M.; Sierwald, Petra; Kiser, Selena B.; Golovatch, Sergei I. (2000). Nomenclator generum et familiarum Diplopodorum II : a list of the genus and family-group names in the class Diplopoda from 1958 through 1999. Sofia, Bulgaria: Pensoft. p. 5. ISBN 954-642-107-3
↑ Hoffman, Richard L. (1980). Classification of the Diplopoda. Geneva, Switzerland: Muséum d'Historie Naturelle. pp. 1–237
↑ Enghoff, H. (1984). «Phylogeny of millipedes – a cladistic analysis». Journal of Zoological Systematics and Evolutionary Research. 22 (1): 8–26. doi:10.1111/j.1439-0469.1984.tb00559.x
↑ Wilson, Heather M.; Shear, William A. (2000). «Microdecemplicida, a new order of minute arthropleurideans (Arthropoda: Myriapoda) from the Devonian of New York State, U.S.A». Transactions of the Royal Society of Edinburgh: Earth Sciences. 90 (4): 351–375. doi:10.1017/S0263593300002674
↑ ab Kraus, O.; Brauckmann, C. (2003). «Fossil giants and surviving dwarfs. Arthropleurida and Pselaphognatha (Atelocerata, Diplopoda): characters, phylogenetic relationships and construction». Verhandlungen des Naturwissenschaftlichen Vereins in Hamburg. 40: 5–50
↑ Kraus, O. (2005). «On the structure and biology of Arthropleura species (Atelocerata, Diplopoda; Upper Carboniferous/Lower Permian)». Verhandlungen des Naturwissenschaftlichen Vereins in Hamburg. 41: 5–23
↑ abcdef Shelley, Rowland M. (1999). «Centipedes and millipedes with emphasis on North American fauna». The Kansas School Naturalist. 45 (3): 1–16
↑ Brewer, Michael S.; Bond, Jason E. (2013). «Ordinal-level phylogenomics of the arthropod class Diplopoda (Millipedes) based on an analysis of 221 nuclear protein-coding loci generated using next-generation sequence analyses». PLoS ONE. 8 (11): e79935. PMC 3827447. PMID 24236165. doi:10.1371/journal.pone.0079935
↑ Blower, John Gordon (1985). Millipedes: Keys and Notes for the Identification of the Species. [S.l.]: Brill Archive. p. 1. ISBN 90-04-07698-0
↑ Brusca, R. C., Brusca, G.J (2007). Invertebrados. [S.l.]: Guanabara-Koogan
↑ ab Minelli, Alessandro; Golovatch, Sergei I. (2001). «Myriapods». In: Levin, Simon A. Encyclopedia of Biodiversity (PDF). [S.l.: s.n.] pp. 291–303. ISBN 0-12-226865-2. Consultado em 11 de julho de 2018. Arquivado do original (PDF) em 21 de fevereiro de 2014
↑ abcdefgh Barnes, Robert D. (1982). Invertebrate Zoology. Philadelphia, PA: Holt-Saunders International. pp. 818–825. ISBN 0-03-056747-5
↑ Marek, Paul E.; Moore, Wendy (2015). «Discovery of a glowing millipede in California and the gradual evolution of bioluminescence in Diplopoda». Proceedings of the National Academy of Sciences. 112 (20): 6419–6424. Bibcode:2015PNAS..112.6419M. doi:10.1073/pnas.1500014112
↑ Lewis, J. G. E. (2008). The Biology of Centipedes A 1ª versão no livro de bolso está digitalizada. ed. Cambridge: Cambridge University Press. pp. 110–111. ISBN 978-0-521-03411-1
↑ Capinera, John L., ed. (2008). «Millipedes». Encyclopedia of Entomology. [S.l.]: Springer. pp. 2395–2397. ISBN 978-1-4020-6242-1
↑ Mesibov, Robert. «Paranota». External Anatomy of Polydesmida. Consultado em 30 de outubro de 2013
↑ abcde Hopkin, Stephen P.; Read, Helen J. (1992). The Biology of Millipedes. [S.l.]: Oxford University Press. ISBN 0-19-857699-4
↑ Marek, P.; Shear, W.; Bond, J. (2012). «A redescription of the leggiest animal, the millipede Illacme plenipes, with notes on its natural history and biogeography (Diplopoda, Siphonophorida, Siphonorhinidae)». ZooKeys. 241 (241): 77–112. PMC 3559107. PMID 23372415. doi:10.3897/zookeys.241.3831
↑ Brusca, R.C., Brusca, G.J (2007). Invertebrados. [S.l.]: Guanabara-Koogan
↑ abcd Blower, J. Gordon (1985). Millipedes: Keys and Notes for the Identification of the Species. London: Published for the Linnean Society of London and the Estuarine and Brackish-Water Sciences Association by E.J. Brill. ISBN 90-04-07698-0
↑ Mesibov, Robert. «Gonopods». External Anatomy of Polydesmida. Consultado em 27 de outubro de 2013
↑ Drago, Leandro; Fusco, Giuseppe; Garollo, Elena; Minelli, Alessandro (2011). «Structural aspects of leg-to-gonopod metamorphosis in male helminthomorph millipedes (Diplopoda)». Frontiers in Zoology. 8 (1): 19. doi:10.1186/1742-9994-8-19
↑ Wesener, Thomas; Köhler, Jörn; Fuchs, Stefan; van den Spiegel, Didier (2011). «How to uncoil your partner—"mating songs" in giant pill-millipedes (Diplopoda: Sphaerotheriida)». Naturwissenschaften. 98 (11): 967–975. Bibcode:2011NW.....98..967W. PMID 21971844. doi:10.1007/s00114-011-0850-8
↑ Enghoff, Henrik; Akkari, Nesrine (2011). «A callipodidan cocoon (Diplopoda, Callipodida, Schizopetalidae)». International Journal of Myriapodology. 5: 49–53. doi:10.3897/ijm.5.1995
↑ Shelley, Rowland M.; Golavatch, Sergei I. (2011). «Atlas of myriapod biogeography. I. Indigenous ordinal and supra-ordinal distributions in the Diplopoda: Perspectives on taxon origins and ages, and a hypothesis on the origin and early evolution of the class». Insecta Mundi. 158: 1–134
↑ ab Golovatch, Sergei I.; Kime, R. Desmond (2009). «Millipede (Diplopoda) distributions: a review.» (PDF). Soil Organisms. 81 (3): 565–597
↑ «Piolho-de-cobra: conheça curiosidades sobre este animal». Natureza e Conservação. Consultado em 10 de agosto de 2017
↑ Adis, Joachim (1986). «An 'aquatic' millipede from a Central Amazonian inundation forest». Oecologia. 68 (3): 347–349. Bibcode:1986Oecol..68..347A. doi:10.1007/BF01036737
↑ Burrows, F. J.; Hales, D. F.; Beattie, A. J. (1994). «Aquatic millipedes in Australia: a biological enigma and a conservation saga». Australian Zoologist. 29 (3–4): 213–216. doi:10.7882/az.1994.007
↑ Barber, A. D., ed. (2013). «World Database of Littoral Myriapoda». World Register of Marine Species. Consultado em 25 de outubro de 2013
↑ Barker, G. M. (2004). «Millipedes (Diplopoda) and Centipedes (Chilopoda) (Myriapoda) as predators of terrestrial gastropods». In: Barker, G. M. Natural Enemies of Terrestrial Molluscs. [S.l.]: CAB International. pp. 405–426. ISBN 978-0-85199-061-3
↑ Weldon, Paul J.; Cranmore, Catherine F.; Chatfield, Jenifer A. (2006). «Prey-rolling behavior of coatis (Nasua spp.) is elicited by benzoquinones from millipedes». Naturwissenschaften. 93 (1): 14–16. Bibcode:2006NW.....93...14W. doi:10.1007/s00114-005-0064-z
↑ Saporito, R. A.; Donnelly, M. A.; Hoffman, R. L.; Garraffo, H. M.; Daly, J. W. (2003). «A siphonotid millipede (Rhinotus) as the source of spiropyrrolizidine oximes of dendrobatid frogs». Journal of Chemical Ecology. 29 (12): 2781–2786. doi:10.1023/B:JOEC.0000008065.28364.a0
↑ Eisner, T.; Eisner, M.; Attygalle, A. B.; Deyrup, M.; Meinwald, J. (1998). «Rendering the inedible edible: circumvention of a millipede's chemical defence by a predaceous beetle larva». Proceedings of the National Academy of Sciences of the United States of America. 95 (3): 1108–13. Bibcode:1998PNAS...95.1108E. PMC 18689. PMID 9448293. doi:10.1073/pnas.95.3.1108
↑ Ito, F. (1998). «Colony composition and specialized predation on millipedes in the enigmatic ponerine ant genus Probolomyrmex (Hymenoptera, Formicidae)» (PDF). Insectes Sociaux. 45 (1): 79–83. doi:10.1007/s000400050070
↑ Herbert, D. G. (2000). «Dining on diplopods: remarkable feeding behaviour in chlamydephorid slugs (Mollusca: Gastropoda)». Journal of Zoology. 251 (1): 1–5. doi:10.1111/j.1469-7998.2000.tb00586.x
↑ Forgie, Shaun A.; Grebennikov, Vasily V.; Scholtz, Clarke H. (2002). «Revision of Sceliages Westwood, a millipede-eating genus of southern African dung beetles (Coleoptera : Scarabaeidae)». Invertebrate Systematics. 16 (6): 931–955. doi:10.1071/IT01025
↑ Larsen, T. H; Lopera, A.; Forsyth, A.; Genier, F. (2009). «From coprophagy to predation: a dung beetle that kills millipedes». Biology Letters. 5 (2): 152–155. PMC 2665820. PMID 19158030. doi:10.1098/rsbl.2008.0654
↑ Forthman, M.; Weirauch, C. (2012). «Toxic associations: a review of the predatory behaviors of millipede assassin bugs (Hemiptera: Reduviidae: Ectrichodiinae)» (PDF). European Journal of Entomology. 109 (2): 147–153. doi:10.14411/eje.2012.019
↑ Animals: The International Wildlife Magazine. [S.l.]: Nigel-Sitwell. 1964. p. 21
↑ Blum, Murray S.; Woodring, J. Porter (1962). «Secretion of benzaldehyde and hydrogen cyanide by the millipede Pachydesmus crassicutis (Wood)». Science. 138 (3539): 512–513. Bibcode:1962Sci...138..512B. PMID 17753947. doi:10.1126/science.138.3539.512
↑ Kuwahara, Yasumasa; Ômura, Hisashi; Tanabe, Tsutomu (2002). «2-Nitroethenylbenzenes as natural products in millipede defense secretions». Naturwissenschaften. 89 (7): 308–310. Bibcode:2002NW.....89..308K. PMID 12216861. doi:10.1007/s00114-002-0328-9
↑ Weldon, Paul J.; Aldich, Jeffrey R.; Klun, Jerome A.; Oliver, James E.; Debboun, Mustapha (2003). «Benzoquinones from millipedes deter mosquitoes and elicit self-anointing in capuchin monkeys (Cebus spp.)». Naturwissenschaften. 90 (7): 301–305. Bibcode:2003NW.....90..301W. PMID 12883771. doi:10.1007/s00114-003-0427-2. Consultado em 13 de julho de 2018. Arquivado do original em 23 de setembro de 2017
↑ Valderrama, Ximena; Robinson, John G.; Attygalle, Athula B.; Eisner, Thomas (2000). «Seasonal anointment with millipedes in a wild primate: a chemical defense against insects». Journal of Chemical Ecology. 26 (12): 2781–2790. doi:10.1023/A:1026489826714
↑ Birkinshaw, Christopher R. (1999). «Use of millipedes by black lemurs to anoint their bodies». Folia Primatologica. 70 (3): 170–171. doi:10.1159/000021691
↑ Roncadori, R. W.; Duffey, S. S.; Blum, M. S. (1985). «Antifungal activity of defensive secretions of certain millipedes». Mycologia. 77 (2): 185–191. JSTOR 10.2307/3793067. doi:10.2307/3793067
↑ Eisner, Thomas; Eisner, Maria; Deyrup, Mark (1996). «Millipede defense: use of detachable bristles to entangle ants» (PDF). Proceedings of the National Academy of Sciences. 93 (20): 10848–10851. Bibcode:1996PNAS...9310848E. PMC 38244. PMID 8855269. doi:10.1073/pnas.93.20.10848
↑ Stoev, Pavel; Lapeva-Gjonova, Albena (2005). «Myriapods from ant nests in Bulgaria (Chilopoda, Diplopoda)» (PDF). Peckiana. 4: 131–142
↑ Farfan, Monica; Klompen, Hans (2012). «Phoretic mite associates of millipedes (Diplopoda, Julidae) in the northern Atlantic region (North America, Europe)». International Journal of Myriapodology. 7: 69–91. doi:10.3897/ijm.7.3064
↑ Swafford, Lynn; Bond, Jason E. (2010). «Failure to cospeciate: an unsorted tale of millipedes and mites». Biological Journal of the Linnean Society. 101 (2): 272–287. doi:10.1111/j.1095-8312.2010.01499.x
↑ Martínez-Torres, Shirley Daniella; Daza, Álvaro Eduardo Flórez; Linares-Castillo, Edgar Leonardo (2011). «Meeting between kingdoms: discovery of a close association between Diplopoda and Bryophyta in a transitional Andean-Pacific forest in Colombia». International Journal of Myriapodology. 6: 29–36. doi:10.3897/ijm.6.2187
↑ Marshall, Michael (22 de setembro de 2011). «Zoologger: Stealth millipede wears living camouflage». New Scientist. Consultado em 26 de junho de 2016
↑ Mason, G.; Thompson, H.; Fergin, P.; Anderson, R. (1994). «Spot diagnosis: the burning millipede». Medical Journal of Australia. 160 (11): 718–726. PMID 8202008
↑ Shpall, S.; Frieden, I. (1991). «Mahogany discoloration of the skin due to the defensive secretion of a millipede». Pediatric Dermatology. 8 (1): 25–27. PMID 1862020. doi:10.1111/j.1525-1470.1991.tb00834.x
↑ Radford, A. (1976). «Giant millipede burns in Papua New Guinea». Papua New Guinea Medical Journal. 18 (3): 138–141. PMID 1065155
↑ Radford, A. (1975). «Millipede burns in man». Tropical and Geographical Medicine. 27 (3): 279–287. PMID 1103388
↑ Hudson, B.; Parsons, G. (1997). «Giant millipede 'burns' and the eye». Transactions of the Royal Society of Tropical Medicine and Hygiene. 91 (2): 183–185. PMID 9196764. doi:10.1016/S0035-9203(97)90217-0
↑ Alagesan, P.; Muthukrishnan, J. (2005). «Bioenergetics of the household pest, Xenobolus carnifex (Fabricius, 1775)» (PDF). Peckiana. 4: 3–14
↑ Enghoff, Henrik; Kebapći, Ümit (2008). «Calyptophyllum longiventre (Verhoeff, 1941) invading houses in Turkey, with the first description of the male (Diplopoda: Julida: Julidae)». Journal of Natural History. 42 (31–32): 2143–2150. doi:10.1080/00222930802196055
↑ Ebregt, E.; Struik, P. C.; Odongo, B.; Abidin, P. E. (2005). «Pest damage in sweet potato, groundnut and maize in north-eastern Uganda with special reference to damage by millipedes (Diplopoda)». NJAS – Wageningen Journal of Life Sciences. 53 (1): 49–69. doi:10.1016/S1573-5214(05)80010-7
↑ Niijima, Keiko (2001). ヤケヤスデ列車を止める [A millipede outbreak (Oxidus gracilis, Koch) stopped trains]. Edaphologia (em Japanese) (68): 43–46. ISSN 0389-1445 [ligação inativa] !CS1 manut: Língua não reconhecida (link)[ligação inativa]
↑ Peckham, Matt (4 de setembro de 2013). «Millipedes – Yes, Millipedes – May Be Responsible for Australian Train Crash». Time Newsfeed. Time Magazine. Consultado em 31 de outubro de 2013
↑ Stoev, Pavel; Zapparoli, Marzio; Golovatch, Sergei; Enghoff, Henrik; Akkari, Nesrine; Barber, Anthony (2010). «Myriapods (Myriapoda). Chapter 7.2. In: Roques et al. (Eds). Alien terrestrial arthropods of Europe». BIORISK – Biodiversity and Ecosystem Risk Assessment. 4: 97–130. doi:10.3897/biorisk.4.51
↑ Lewbart, Gregory A., ed. (20 de setembro de 2011). Invertebrate Medicine 2nd ed. [S.l.]: Wiley-Blackwell. p. 255. ISBN 978-0-470-96078-3
↑ abc Costa Neto, Eraldo M. (2007). «The perception of Diplopoda (Arthropoda, Myriapoda) by the inhabitants of the county of Pedra Branca, Santa Teresinha, Bahia, Brazil». Acta Biológica Colombiana. 12 (2): 123–134
↑ Lawal, O. A.; Banjo, A. D. (2007). «Survey for the usage of arthropods in traditional medicine in southwestern Nigeria» (PDF). Journal of Entomology. 4 (2): 104–112. doi:10.3923/je.2007.104.112
↑ Negi, C. S.; Palyal, V. S. (2007). «Traditional uses of animal and animal products in medicine and rituals by the Shoka tribes of district Pithoragarh, Uttaranchal, India» (PDF). Studies on Ethno-Medicine. 1 (1): 47–54
↑ Jiang, T. L.; Feng, G. W.; Shen, J. H.; Li, L. F.; Fu, X. Q. (1981). «Observation of the effect of Spirobolus bungii extract on cancer cells». Journal of Traditional Chinese Medicine. 1 (1): 34–8. PMID 6926686
↑ Enghoff, Henrik; Manno, Nicola; Tchibozo, Sévérin; List, Manuela; Schwarzinger, Bettina; Schoefberger, Wolfgang; Schwarzinger, Clemens; Paoletti, Maurizio G. (2014). «Millipedes as food for humans: their nutritional and possible antimalarial value: a first report». Evidence-Based Complementary and Alternative Medicine. 2014: 1–9. doi:10.1155/2014/651768
↑ «Canada: Money in muskeg?». Reed Business Information. New Scientist: 198–199. 25 de abril de 1963. ISSN 0262-4079
↑ Avirovik, Dragan; Butenhoff, Bryan; Priya, Shashank (2014). «Millipede-inspired locomotion through novel U-shaped piezoelectric motors». Smart Materials and Structures. 23 (3): 037001. Bibcode:2014SMaS...23c7001A. doi:10.1088/0964-1726/23/3/037001
↑ Wakimoto, Shuichi; Suzumori, Koichi; Kanda, Takefumi (2006). «A bio-mimetic amphibious soft cord robot». Nihon Kikai Gakkai Ronbunshu, C Hen/Transactions of the Japan Society of Mechanical Engineers, Part C (em Japanese e English). 72 (2): 471–477 !CS1 manut: Língua não reconhecida (link)
↑ Beattie, Andrew; Ehrlich, Paul (2001). Wild Solutions: How Biodiversity is Money in the Bank 2nd ed. New Haven: Yale University Press. pp. 192–194. ISBN 978-0-300-10506-3
Ver também |
- Miriápodes
- Quilópodes
Ligações externas |
- SysMyr, uma base de dados da taxonomia dos miriápodes
- British Myriapod & Isopod Group
| Controle de autoridade |
|
|---|


